核酸・タンパク質の構造・機能の特性を生かした分析技術・創薬開発に関する研究
核酸・タンパク質などの生体分子は、生命の活動を司る重要な分子です。これらの分子の構造や分子認識能を理解し、特性を生かすことで、新しい分析技術や医薬品の開発に結び付けることが可能です。また、生体分子の機能を凌駕する「人工」分子が開発できれば、付加価値の高い機能性材料や医薬品の創出が期待できます。これらの観点から、生命分析化学研究室では、新規核酸医薬品デザイン、核酸医薬品の課題克服、生体分子を検出する機能性核酸・人工タンパク質の開発、などの研究に取り組みます。
核酸医薬品は、疾患標的を遺伝子発現レベルで制御する新しい機序の医薬品です。低分子化合物ではアプローチが難しいとされてきた疾患標的に対しても有効な治療薬が開発できる可能性が高く、近年活発に研究が進められています。私たちの研究室では、有効性が高く、副作用が低く、安全性の高い核酸医薬品の開発のための技術開発を目指します。
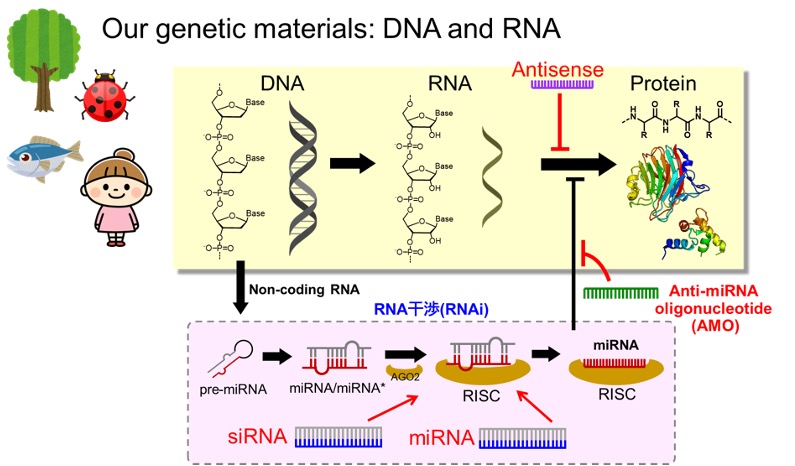
これまで名古屋大学大学院工学研究科浅沼研究室で開発された人工核酸SNA、L-aTNAを用いて、核酸医薬品の基盤設計を開発してきました。本技術をベースに、核酸医薬品に求められる機能開発や課題解決に取り組み、高性能な核酸医薬品開発へと発展を目指します。
《参考文献》
J. Org. Chem., 2023, 88, 2, 796–804
Chem. Asian J., 2020, 15, 1266-1271
ChemBioChem, 2017, 18, 1917-1922
ChemBioChem, 2014, 15, 2549-2555
核酸やタンパク質の優れた性質を利用することで、生命現象の理解を促す分子ツールや、診断に役立つ分析手法の開発を行います。
例えば、バイオマーカーとなる生体分子や低分子を特異的に検出する手法が開発できれば、疾病の診断、病態の解析、薬物検査が可能となります。また、興味の対象となっている生体分子の細胞内動態を可視化する、また、生体内での相互作用を抽出することができれば、対象とする分子が機能する仕組みを明らかにすることができます。このように、社会に役立つ、また、生命科学に役立つバイオテクノロジーツールの開発に挑みます。
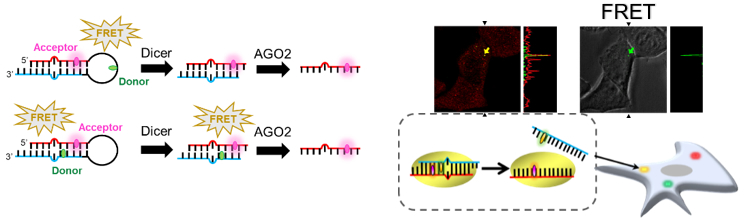
《参考文献》
Sensors, 2021, 21, 1785.
Int. J. Mol. Sci., 2020, 21, 5218
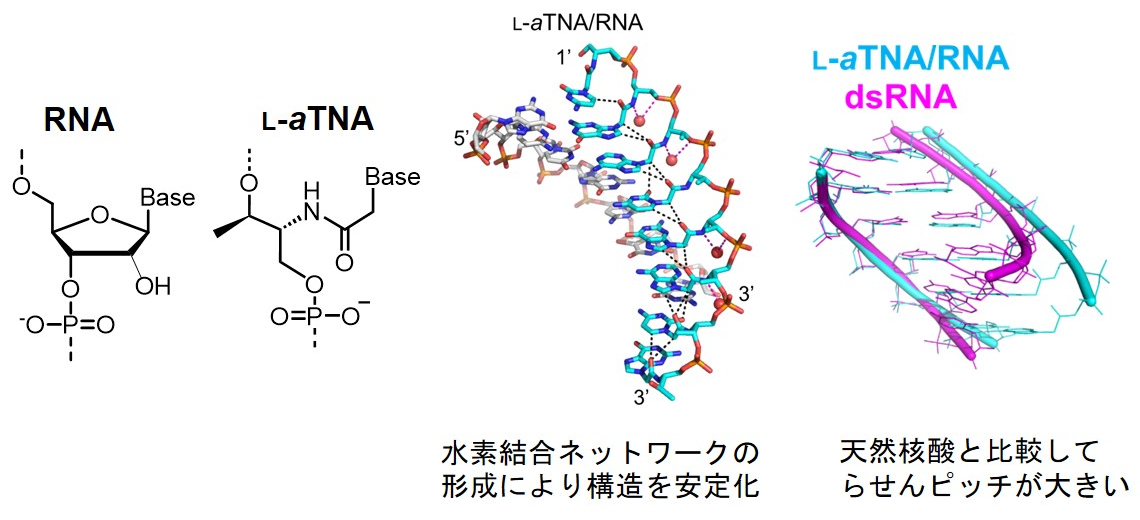
地球上の生命は、DNAとRNAとタンパク質を要素とするセントラルドグマに支配されています。いずれの生命体においても、DNAとRNAは遺伝情報の保存分子として、タンパク質は生命機能の発動分子として役割を果たしていますが、一方で核酸化学やタンパク質科学の進展により、天然にはない核酸やタンパク質が開発されており、ときには天然を超える性能をもつものが開発されています。なぜ地球生命はDNA・RNA・タンパク質を生命分子として選択したのか?という本質的な問いに、人工核酸の核酸認識能・核酸構造の理解・人工タンパク質の設計、そしてこれらの天然分子との比較を通じて、DNAおよびRNAが生命に選択された謎を紐解き、生命の起源あるいは人工生命の創出に対する新たな視点を得たいと考えています。
《参考文献》
Commun. Chem., 2020, 3, 156
〒658-8558
兵庫県神戸市東灘区本山北町4丁目19番1号
TEL 078-441-7548 FAX 078-441-7550
Copyright© Kobe Pharmaceutical University. All rights reserved.